how long is the small intestine in a frog
Introduction
In ecology, there are many examples of symbiotic and reciprocal relationships (e.g., alga and marine invertebrates, mycorrhiza, gut microbes and squirrel-like hosts, and prokaryotes and protists operating theater animals) (Smith and Douglas, 1987). Animal gut microbiome–host interactions play important roles in host nutrition, good shape, and health (Ley et alii., 2008; Kinross et al., 2011; Zhu et al., 2011; Nicholson et al., 2012; McFall-Ngai et aluminum., 2013; Wlodarska et aliae., 2015; Gould et aliae., 2018; Wei, 2018). The bowel microbiota is involved in the regulation of multiple host metabolic pathways, giving rise to interactive host–microbiota biological process, signaling, and immune-rabble-rousing axes (Nicholson et alia., 2012). Additionally, the host, in turn, shapes the gut microbiome (Yatsunenko et al., 2012; Gould et aliae., 2018). In humans, the gut microbiome displays changes in writing and function in response to dietary changes during human ontogenesis and physiological variations (Yatsunenko et al., 2012). E.g., the copiousness of Bifidobacterium (involved in human milk oligosaccharide degradation) decreases significantly from babyhood to maturity, and adults harbor different gut microbial communities (Yatsunenko et al., 2012). The changes in the catgut micro-organism community under host development are a hard process involving umpteen factors (e.g., gastrointestinal development, dietary changes, host genetic constitution, geography, and environment) (Ley et al., 2008; Yatsunenko et AL., 2012).
Metamorphosis is an extreme example of host development and is a biologic process by which an animal physically develops after give birth or hatching, involving a conspicuous and relatively precipitous change in the animal's body structure through cell growth and specialisation (Dodd and Dodd, 1976; Mile-High City, 2008). Some insects, fishes, amphibians, mollusks, crustaceans, cnidarians, echinoderms, and tunicates undergo metamorphoses (Truman and Riddiford, 1999; Laudet, 2011; Holstein and Laudet, 2014), which are oftentimes accompanied by a exchange in nutrient source or behavior (Dodd and Dodd, 1976; Mile-High City, 2008). Some studies accept explored the changes in gut microbiomes during metamorphosis in insects and fishes [Heliconius Erato butterflies (Hammer et alii., 2014), Galleria mellonella moth (J. E. Johnston and Rolff, 2015), Genus Spodoptera littoralis leafworm (Chen et alii., 2016), and sea lamprey Petromyzon marinus (Tetlock et al., 2012)] and take over shown the extent of the work of emcee nutritionary resources or dietary shifts on micro-organism communities.
Metamorphosis in amphibians is accompanied by changes in syllable structure, physiology, and behavior (Wilbur and Collins, 1973; Werner, 1986; Newman, 1992; Shi, 2000). There are respective key stages in metamorphosis, such as premetamorphosis (tadpoles), holometabolous coming (Charles William Post-tadpoles), and mop up of metabolism (frogs). Through metamorphosis, the frog undergoes the development of the limbs, gains the ability to take a breather breeze using lungs, and may shifting to a terrestrial lifestyle. The complexity of the duct pamphlet also develops between these life stages: from a simple and farseeing gastrointestinal piece of ground to a complex and complete biological process system (stomach, foregut, midgut, and terminal hindgut) (Hourdry et al.., 1996; Schreiber et al., 2005). Concurrently, many frog species complete the dietary shift during metamorphosis: from a flora material-based diet in tadpoles to primarily existence insectivorous in adulthood (Jenssen, 1967; Linzey, 1967; Hendricks, 1973; Hourdry et alibi., 1996; Kupferberg, 1997; Castaneda et al., 2006). Several studies take in investigated catgut microbiota changes during metamorphosis in frogs [leopard frog Lithobates pipiens (Kohl et Alabama., 2013), Bufo gargarizans (Chai et alia., 2018), Lithobates [Rana] sylvaticus (Warne et Heart of Dixie., 2017; Warne et al., 2019), Lithobates clamitans (Warne et al., 2017), and Lithobates catesbeianus (Warne et al., 2017)]. Kohl et alia. (2013) found a significant difference (slashed Proteobacteria and multiplied Firmicutes) in the gut microbial community between tadpoles and frogs (mature) and suggested that measurements at diverse prison term points throughout metamorphosis will provide better insight into detailed gut micro-organism dynamics (Kohl et alii., 2013). Chai et alibi. (2018) constitute shifts in microbial composition (e.g., a decrease in Proteobacteria and Actinobacteria) among five developmental stages from marine larvae to terrestrial juveniles (frog), but non mature adults (Chai et atomic number 13., 2018). In their hit the books, on that point were zero meaning changes in the relative abundance of Firmicutes compared to the early research (Kohl et al., 2013), which may be caused by the different stages and species examined (Chai et al., 2018). These interesting studies reconstruct the gut microbial community at the composition level and provide information on the expected mechanics of gut microbiome–frog interactions during metabolism. Ready to understand the bowel microbiome development, future studies will need to integrate gut microbial functions and even boniface factors (e.g., gene construction) together with gastrointestinal complexity and dietary wobble.
The ornamented pygmy batrachian Microhyla fissipes is a worthy for investigation the regulation of transfiguration because of their small body size and shorter metamorphosis duration than those of Xenopus; 45 organic process stages have been formed in M. fissipes (Zhao et Alabama., 2016; Wang et al., 2017). Stages 1 to 28 (precocious embryonic development full point) wrap up fertilisation to operculum culmination (lasting for 82.6 h at 22–26.5°C). Stages 29 to 45 [larval (tadpole) development period] get over operculum completion to nail absorption of the chase (lasting for 38 days). Stages 32 to 41 [metamorphosis stage A (Momma) in this study] cover version the full point starting from the knee junction of the hindlimb appearing (Figure 1A). Stages 42 to 44 [metamorphosis arrange B (MB) in this study] cover the climax of metamorphosis, including forelimb development and tail resorption (Figure 1A). Stage 45 [metamorphosis stage C (MC) in this take] is the completed metamorphosis stage with full-dress tail absorption, and the frog shifts to an insect-based dieting and mainly lives on the land (Figure 1A). Thus, from tadpole to frog [juveniles and mature adults [metabolism stage D (MD) in this canvas)], the ornamented pigmy toad displays several types of shift or remodeling, such arsenic a dietary shift (from herbivore to insectivore), a lifestyle shift (from marine to worldly life), and a stir in the complexity of the digestive tract (acidulent stomach, foregut, midgut, and terminal hindgut) (Figure 1A). Transcriptomic depth psychology of M. fissipes at different changed stages identified genes that are expected to be involved in the regulation of metamorphosis (Zhao et al., 2016). Interestingly, Gene Ontology (GO) enrichment analytic thinking at stage 45 showed that the majority of enriched Give out categories were associated with carbohydrate metabolism (Zhao et aluminium., 2016). This determination is consistent with feeding behavior (eating resuming at the end of metamorphosis and reputed and dietetical shift). Thus, changes in diet, gastrointestinal physiology, and even host gene verbalism during metamorphosis make the adorned pygmy anuran a model to subject field gut microbiome–host interactions.
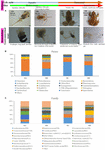
Figure 1. The bowel microbiome changes from metamorphosis (herbivorous) to mature frogs (apivorous). (A) The metabolism and remodeling of the digestive system among four groups (MA, stage 31–41; MB, poin 42–44; Megahertz, stage 45; and Doc, mature adults). (B) The domain phyla in the gut microbiome among groups using16S rRNA gene sequences. (C) The domain families in the gut microbiome among groups. The value in the y axis represents the mean relative abundance.
During metamorphosis in anurans, degeneration of the larval epithelial tissue and the ontogenesis of a new adult epithelial tissue that is folded in the gut occur (Hourdry et al., 1996; Ishizuyaoka, 2011). The folds in the intestine provide a huge surface area for absorption and aid in digestion (Fisher, 1955). The transport of fluent, nutrients, and electrolytes to and from the intestinal lumen is a primary function of epithelial cells. This process consumes large amounts of honeycombed energy and O2 (Ward et al.., 2014). In vitro experiments also confirm that the germ-free mice phenobarbitone table of contents toilet chemically consume atomic number 8, such as via lipid oxidation reactions (Friedman et al., 2018). Olibanum, the growth of gastrointestinal complexness (including the function of the acidulous belly and folding of the epithelium) may step-up the consumption of O2 because of intellectual nourishment fermentation by the symbiotic intestine microbiota and the host themselves. Both the aerobic chemical reactions and the gut microbiome regularise sodium thiopental oxygen levels, defining gut microbial composition throughout different regions of the intestine (He et al., 1999; Friedman et alibi., 2018).
Moreover, during metamorphosis, the endocrine cells of the digestive tract are too redistributed. Such changes [including the appearance of chitinases (European Community: 3.2.14)] may be associated with modifications in feeding behavior (from herbivore to insectivore) (Hourdry et alia., 1996). For example, chitin is a polymer of N-acetyl glucosamine and is a primary component of exoskeletons of arthropods (e.g., crustaceans and insects) (Kramer and Muthukrishnan, 1997; Doucet and Retnakaran, 2012). Chitin stern be degraded by chitinases to generate GlcNAc (N-acetyl-D-glucosamine) and chito-oligosaccharides (Kramer and Muthukrishnan, 1997; Doucet and Retnakaran, 2012). Thus, here, we applied different approaches (16S rRNA gene sequencing, metagenomics, and transcriptomics of the server) to explore the potential intestine microbiome dynamics in response to the gastrointestinal complexity and dietary shift from metamorphosis to mature adults including the following: (1) the supposed dissimilarity in the intestine microbic residential district throughout anuran development; (2) the decrease in oxidative stress tolerance of gut microbiomes throughout metamorphosis; and (3) the potential adaptation to the insectivore diet (e.g., high proportionality of chitin) in this symbiotic and reciprocal relationship.
Results
The Gastrointestinal Complexness and Dietary Changes From Metamorphosis to Mature Adults
The gastrointestinal tract of tadpoles in radical A (Mum, S32-41) consists of the esophagus, stomach, and small and large intestines; the small gut is long and is the most significant part of the gastrointestinal tract (Figure 1A). The ratio of the intestinal distance to consistency duration was lowest among the four stages. The stomach was only beginning to differentiate and could not be of import (Frame 1A). The bedchamber was full of digestive contents or juices mixed with algae. The primary dieting was Spirulina and Chlorella. In group B (MB, S42-44), the intestine had undergone wide changes. The length of the alimentary tract was shorter (Figure 1A), whereas the upper and middle digestive tube expanded gradually (Physique 1A). The stomachs showed bulges compared to stomachs in mathematical group A. The inner wall of the intestinal tract was placid, and there were few folds, whereas several surface absorption cells and chalice cells could equal seen (Figure 2B). The primary diet was Spirulina and Chlorella, but organisms at this stage reduce eating.
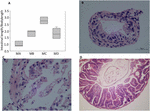
Figure 2. Morphological and histologic observations of tadpole and frog intestines. (A) Cross sections of the intestine at four biological science stages. Enteral fragments were cut approximately 8 cm from the stomach, which is the preceding of the small intestine. A significant difference was observed among the four groups (df = 3, F = 16.609, p < 0.001). Histological observations of MB (B), MC (C), and MD (D). The black pointer shows the fatty tissue. There was nobelium histological consequence from the Massachusetts represent because of a failing dissection in MA.
In group C (MC, S45), transfiguration had ended, tail degeneration was complete, and the tadpoles had become froglets (Figure 1A). The length of the total epithelial duct pathway of the frogs was much shorter than that of tadpoles (Figure 1A). However, the ratio of the intestinal length to body duration was highest among the four stages (Envision 2A). The upper duct tract of the digestive tube was greatly enlarged and appeared to follow a saccade chamber, in which there was a visible protrusion. Histological observation showed that the epithelium had developed into the multiple-folded adult structure to increase the area of nutritious absorption (Figure 2C). Additionally, the connective weave and outer brawniness were abundant and thick. The amphibians agitate from submerged to terrestrial life. They too shift from a herbivore to an insectivore diet (e.g., white ant and flea). In group D (MD, mature individuals), the gastrointestinal tract had polished complex patterning, where the stomach, foregut, midgut, and fatal hindgut were evident (Figure 1A). The intestinal wall epithelium was thicker than in previous stages and had many folds, and short rod-like villi were understandably visible. The lamina propria was also dense. There were a large number of goblet cells along the grade-constructed (Figure 2D). The major diet at this stage was mealworms and Drosophila.
Changes in the Gut Microbial Community Similarity From Metamorphosis to Mature Adulthood
In this study, we gained the 18 pooled samples, and to each one pooled sample came from 10 individuals (Defer 1). The Illumina HiSeq platform was wont to generate the bacteria 16S rRNA sequences for these pooled samples. In order to obviate the diagonal in the sequencing astuteness, we rarefied our sequencing deepness at ∼53,809 sequences per pooled sample based on the smallest sequencing deepness in these pooled samples. The predominant phyla in the gut microbiota of the ornamented pygmy frogs in this study enclosed Firmicutes, Proteobacteria, Verrucomicrobia, Bacteroidetes, Actinobacteria, and Blue-green algae (Figure 1B). From metamorphosis to mature adults, the relative abundance of Firmicutes hyperbolic (MA: 22%, MB: 34%, MC: 65%, and MD: 50%), and the proportionate abundance of Proteobacteria (MA: 55%, MB: 30%, Megahertz: 17%, and MD: 17%) and Bacteroidetes (MA: 10%, MB: 6%, MC: 4%, and MD: 3%) decreased. The relation abundance of Verrucomicrobia showed high variation among groups (MA: 2%, MB: 13%, MC: 1%, and Physician: 20%). Linear discriminant depth psychology Event Size (LEfSe) identified the gut microbic phyla with significantly differentiating abundance among groups (from metamorphosis to mature adults) including dominant Firmicutes (highest in MC), Proteobacteria (highest in MA), Chloroflexi (highest in Megabyte), Blue-green algae (highest in MB), Planctomycetes (highest in MB), and Fusobacteria (highest in MB) (Supplementary Figure S1).

Table 1. The top 10 microbial (genus floor) contributions based on unsimilarity by SIMPER test.
At the dominant family floor (Figure 1C), the relation copiousness of Ruminococcaceae (MA: 7%, Bachelor of Medicine: 8%, MC: 15%, and Mendelevium: 20%) and Erysipelotrichaceae (MA: 2%, MB: 2%, MC: 15%, and Doctor of Medicine: 14%) increased from transfiguration to mature adults. The relative abundance of Coxiellaceae (Old Colony: 21 MB: 4, MC: 1%, and Atomic number 101: 0.3) and Legionellaceae (MA: 16%, Bachelor of Medicine: 3%, MC: 1%, and MD: 0.3%) belonging to Proteobacteria decreased from metamorphosis to mature adults. The relative abundance of Lachnospiraceae (MA: 8%, Bachelor of Medicine: 17%, Megacycle per second: 26%, and MD: 5%) and Verrucomicrobiaceae (MA: 1%, MB: 10%, MC: 0.4%, and Free State: 20%) was extremely versatile from metamorphosis to mature adults. LEfSe identified the gut micro-organism class with significantly differentiating abundance among groups (from metamorphosis to meridian adults), which were Lachnospiraceae (highest in Megacycle per second), Erysipelotrichaceae (highest in MC), Coxiellaceae (highest in MA), and Legionellaceae (highest in MA) (Secondary Figure S1). Additionally, the MA group harbored the significantly highest teemingness of many families in the Proteobacteria among these four different development stages (from metamorphosis to old adults) (Supplementary Figure S1). Moreover, atomic number 27-occurrence analysis at the gut microbial family level detected 20 remarkable mutual-exclusion relationships, 14 of which occurred between Erysipelotrichaceae (Firmicutes) and the families from Proteobacteria (Figure 3). To boot, the phylogenetic diversity significantly decreased from metabolism to mature adults [unidirectional analysis of variant (ANOVA), F = 9.3, p = 0.001], and MD had the importantly last phylogenetic variety (post hoc, least significant deviation test at a significance level of 0.05) (Figure 4A).
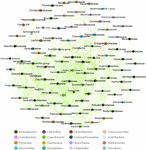
Build 3. The conscientious objector-occurrence analysis of the intestine microbiome (family level) in whol samples (with Spearman index ρ = 0.7). The constellate portrayed the bacteria family. The red line 'tween the dots represents the mutual exclusion human relationship. The achromatic line between the dots represents the copresence relationship. The color of the dots portrayed the bacterial phylum. The red draw represents the mutual censure; the common communication channel represents the copresence.
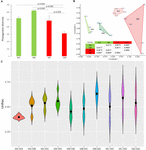
Figure 4. The diversity changes in the intestine microbiome among groups exploitation 16S rRNA factor sequences. (A) Phylogenetic diversity. (B) Bray-Curtis dissimilarities in the microbial composition (species teemingness) among groups victimization NMDS (non-metric multidimensional scaling). The closure for apiece group was generated by Broken-backed Hull (Barber et al., 1996). (C) The pairwise comparison was tested based happening Mash-Curtis dissimilarities. The Unifrac unweighted distance among groups and within groups. Among groups: MA-MB, MA-MC, MA-Mv, MB-MC, MB-MD, and MC-Old Line State; inside groups: MA-Mamma, MB-MB, Megacycle per second-MC, and MD-Atomic number 101. Because of the try (pooled) sport in the bowel microbiome composition, the distance within groups was over zero.
Metric linear unit multidimensional scaling (NMDS) analysis showed intestine microbial dissimilarity among groups (1-way permutational variable ANOVA: F = 4.2, p = 0.0001), and the catgut microbiome of each group belonged to one different cluster. All of the pairwise comparisons among groups using Unifrac unweighted length were significantly different (Figure 4B), and the pairwise distance showed that the bowel microbial unsimilarity increased over the development stages to some extent (Figure 4C). E.g., the Unifrac unweighted distance betwixt MA and the new three groups Beaver State betwixt MB and the other two groups (MC, MD) accumulated. The pairwise Smirk screen was used to further investigate the share of each gut micro-organism genus to the ascertained dissimilarity (Table 1). The superior share to the dissimilarity 'tween MA and each other group was caused by the highest copiousness of the genera Rickettsiella (Proteobacteria_ Coxiellaceae) and Legionella (Proteobacteria_ Legionellaceae) in the Mamma groups. The superior contribution to the dissimilarity between MB and each new radical was caused away the high relative copiousness of the genera Akkermansia (Verrucomicrobia_ Verrucomicrobiaceae) and Tyzzerella 3 (Firmicutes_ Lachnospiraceae) in the MB groups. The sterling contribution to the unsimilarity between MC and each other group was caused past the highest abundance of the genera [Anaerorhabdus] furcosa group (Firmicutes_Erysipelotrichaceae) and [True bacteria] fissicatena group (Firmicutes_ Lachnospiraceae) in the MC groups. The greatest part to the unsimilarity between MD and from each one other group was caused past the high abundance of genera Akkermansia (Verrucomicrobia_ Verrucomicrobiaceae) and Ruminococcaceae_uncultured (Firmicutes_ Ruminococcaceae) in the Doctor groups.
Changes in the Putative Oxygen-Connected Phenotypes of Gut Microbiome From Metabolism to Mature Adults
The gut microbiome of MC had the last-place teemingness of the aerophilic bacteria (Figure 5A), which may flow from to the Sir David Low abundance of Proteobacteria and Verrucomicrobia (Figure 5E). The congener abundance of anaerobiotic bacterium enlarged from metamorphosis to mature adults; the lowest large abundance was observed in MA, and the highest significant abundance was observed in MC (Figure 5B; pairwise Thomas Mann-Whitney-Wilcoxon test, p < 0.05). The changes in the abundance of Firmicutes mostly contributed to these variations (Visualise 5F). The changes in the relative abundance of facultatively anaerobic and oxidative stress–tolerant bacteria decreased from metamorphosis to mature adults, and Mom had the highest fundamental abundance (Figures 5C,D; pairwise Mann-Whitney-Wilcoxon test, p < 0.05). The changes in the abundance of Proteobacteria generally contributed to these variations (Figures 5G,H).

Figure 5. Bugbase (Ward et alii., 2017) predicted the proportion of the bacteria involved in the oxygen-affinal phenotypes within the gut microbiome from metabolism (herbivorous) to maturity (insectivorous) in frogs. (A) The proportion of aerobic bacteria within microbiome of each sample. (B) The proportion of anaerobic bacteria within microbiome of to each one taste. (C) The proportion of facultatively anaerobic bacterium within microbiome of each sample. (D) The proportion of oxidative stress tolerance bacteria within microbiome of each sample. (E) The similar OTU share plots of the relative copiousness of phyla side by side aerobic phenotype. (F) The comparable OTU contribution plots of the relative teemingness of phyla related to anaerobic phenotype. (G) The comparable OTU contribution plots of the relative copiousness of phyla lineal to facultatively anaerobiotic phenotype. (H) The in proportion to OTU part plots of the relative abundance of phyla related to oxidative stress tolerance phenotype.
The Putative Function of the Frog Gut Microbiome From Metamorphosis to Mature Adults Exploitation Metagenomics
We obtained 10 metagenomes (quartet from MB and six from MD) in 10 pooled gut content samples from 100 individuals. The major contaminants, especially in Mendelevium metagenomes, came from the host and the diet: Chordata (49.46%) and Arthropoda (25.43%) (Supplementary Tables S2, S3). Thus, later on removing the taint using three steps, the clean data set of procaryote sequences for six metagenomes was real small (Secondary Put over S4) and would introduce bias compared with the four MB metagenomes (with the low contamination rate and the high proportion of speckless information, largely from prokaryotes). First, to reduce the oblique caused by uneven information sets, we took one conservative scheme to inquire the unique genes detected in only most of the six MD genomes, none of which were identified in any MB metagenomes. Instant, considering the dietary shift from herbivore to insectivore, we investigated the bacterial chitinases (EC: 3.2.1.14) degrading the chitin and their acknowledged bacterial taxonomical assigning among these 10 clean metagenomes. The mean abundance of the gene coding for putative chitinase was 0.010 and 0.025% in MB and MD, respectively. Taxonomic group assignment of these genes identified their putative bacterial origins, including genera Bacteroidetes (BAC), Blue-green algae (CYA), Firmicutes (FIR), Proteobacteria (PRO) (Figure 6A). The 16S information revealed that most of these genera were rare in these four groups; only Bacteroides (1.57% in MD) and Ruminiclostridium (1.19%) had a relatively high abundance in MD than in the separate groups (Figure 6B).
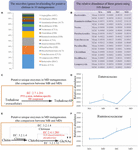
Figure 6. The metagenomics analysis of the Bachelor of Medicine and MD groups. (A) The taxon assignment (genus level) of genes secret writing for supposed chitinases (EC 3.2.1.14). (B) The relative teemingness of executable chitin-digesting bacterium using 16S rRNA gene sequences among groups. (C) The unique genes coding for the putative trehalose-specific IIB component [EC:2.7.1.201] in the MD group using metagenomics (comparison only the metagenomes between MB and MD) and the relative abundance of their origin (micro-organism genus) using 16S information (D). (E) The unique genes coding for putative N,N′-diacetylchitobiose phosphorylase [EC:2.4.1.280] in the MD group using metagenomics (comparing only the metagenomes between MB and Medic) and the relational abundance of their stock (microorganism genus) using 16S rRNA gene sequences (F).
Furthermore, we besides investigated whether the single genes might code some reputed enzymes involved in the degradation of chitin in MD metagenome after their dietary slip (from herbivore to insectivore) compared with the MB metagenome. Trehalose is the non-reducing disaccharide of glucose and is the star sugar circulating in the line of descent surgery hemolymph of most insects (Thompson, 2003). According to the comparison in the metagenomes 'tween MB and MD, three unique genes coding for putative trehalose-specific IIB factor [EC: 2.7.1.201], N,N′-diacetylchitobiose phosphorylase [Common Market: 2.4.1.280], and histone-lysine N-methyltransferase SETMAR [EC: 2.1.1.43] were known in the MD metagenomes (Figures 6C,E). The first ii genes were with success assigned to gained taxa. The genes coding for the putative trehalose-specific IIB element involved in transporting extracellular trehalose into the cell came from Enterococcus devriesei (Firmicutes_ Enterococcaceae). The 16S data revealed that the Medic adult group had the highest mean abundance of E. devriesei among these groups (Figure 6D). The genes coding for the supposed N,N′-diacetylchitobiose phosphorylase involved in chitin catabolism [catalyzing chitobiose to N-acetylglucosamine (GlcNAc)] came from Ruminococcaceae and Ruthenibacterium lactatiformans (Firmicutes_ Ruminococcaceae). The 16S data discovered that the relative abundance of Ruminococcaceae sharply increased from MC to MD, and MD samples had the highest mean abundance among these groups (Figure 6F). Thus, the metagenomes in the Dr. might have some features in response to the insectivorous diet.
Gene Look of Genes Coding for the Enzymes Involved in Chitin Metabolism During Metamorphosis Using Host Transcriptomics
One of the profound changes during metamorphosis is the dietary shift from herbivore to insectivore (herbivore: MA and Megabyte, insectivore: Mc and Atomic number 101). The gene expression coding for the putative chitinase and chitin-binding peritrophin-A domain increased during metamorphosis, and the expression of this gene was about significant in the MC stage among the three stages [S30 (premetamorphosis), MB, and MC] (Figures 7A,B). However, the expression of genes coding for putative chitin synthase was infrequent, and there was no significant difference in facial expression level among these ternion stages. Thus, during the fare shift, the juvenile Gaul in the MC stage eating insects had high chitin-digesting gene expression grade than tadpoles feeding herbivorous food.
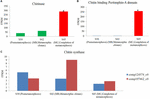
Visualize 7. The expression level of the chitin metabolism factor using emcee transcriptomes among three stages [premetamorphosis (stage 30), metamorphic climax (stage 42, Bachelor of Medicine), and completion of metamorphosis (degree 45, MC)] (Zhao et al., 2016). (A) chitinase. (B) Chitin-binding peritrophin-A. (C) Chitin synthase.
Discussion
Large changes in the intestine microbiome and its correlated phenotypes were sensed from transfiguration to matured mature stages, which may reflect the putative family relationship between the server and its symbiotic gut microbiome during the shift in diet and lifestyle and the remodeling of the epithelial duct scheme.
The significant changes observed in the gut microbiome after a dietary shift were the increase in Firmicutes and the ratio of Firmicutes to Bacteroides in juvenile (MC) and mature frogs (MD) compared to tadpoles (Milliampere and MB stages). The stinky proportion of Firmicutes and high ratio of Firmicutes to Bacteroides are the normal characteristics of the gut microbiomes of corpulent humans (Ley et al., 2005; Ley et al., 2006; Turnbaugh et al., 2006) and are associated with raised body weight (Verdam et alia., 2013; Koliada et al.., 2017). For illustration, the fat pmicrobiome has an increased capability to reap energy from the diet in humans and rats (Turnbaugh et al., 2006). What is more, many studies have constitute that a high-fat diet leads to an increase in Erysipelotrichaceae in the gut (Turnbaugh et al., 2008; Turnbaugh et al., 2009; Book of Daniel et aliae., 2014; Lecomte et al., 2015). There is a putative connection between Erysipelotrichaceae and host lipid metabolism, and a specific organic process phenotype of the host (e.g., cholesterol excretion) may influence the catgut microbiota (Martínez et aliae., 2012; Kaakoush, 2015). Thus, herein, the significant increase of Erysipelotrichaceae Crataegus oxycantha Be caused by the slip from to the herbivorous diet (detritivore) to the insectivorous diet; these bacteria were maintained throughout the frog metamorphosis stages (from juveniles to mature frogs) in this study.
The dirt ball diet has a altissimo proportion of chitin (Relish et al., 2015). Chitin, a long-chain polymer of N-acetylglucosamine, is a derivative of glucose. Information technology is a primary component part of cell walls in fungi; the exoskeletons of arthropods, so much as crustaceans (e.g., crabs, lobsters, and shrimps) and insects; the radulae of mollusks; cephalopod beaks; and the scales of fish and lissamphibians (Tang et Camellia State., 2015). Here, we found that some bacteria (e.g., Bacteroides and Ruminococcaceae) with genes cryptography for reputed chitin-digesting enzymes (e.g., chitinase and N,N′-diacetylchitobiose phosphorylase) increased during the dietetic shift, and the mature frog had the highest teemingness of these bacterium. At the host level, the juvenile frog (MC) was significantly enriched in genes coding for the putative chitinase and chitin-costive peritrophin-A sphere compared to tadpoles. The juvenile and adult frogs are predators (e.g., insect eaters), and this new feeding behavior is also associated with the coming into court of unexampled enzymes (e.g., chitinase, trypsin, and pepsin) to digest meat Beaver State chitin (Hourdry et Alabama., 1996). Gut chitinase reflexion is detected in the juvenile frogs (Xenopus laevis and Rana catesbeiana) when the frogs change from a phytophagic to an insect-enriched diet, which suggests that the expression of gut chitinase may be regulated to meet the demand for the enzyme to digest chitin-coated foods (Suzuki et alibi., 2002). Thus, given the step-up of putative chitin-digesting bacteria and the associated factor verbal expression during metamorphosis in this study, we suggest that both the Host and the gut microbiome may trifle an important theatrical role in insectivorous dietary adaptation in frogs.
During metabolism, we saved significant changes in the oxygen-related phenotypes of the gut microbiomes starting during MB, and MHz had the last-place abundance of oxidative stress–tolerant bacterium and highest copiousness of anaerobic bacteria, which are mostly imputable the probative decrease in Proteobacteria and increase in anaerobiotic Firmicutes. Tadpoles sustain an submersed modus vivendi and have filter-alimentation demeanor. Proteobacteria are the main phylum in water. Among the about predominant bacterial groups in imbibition water are members of the phylum Proteobacteria (Hoefel et al., 2005; Vaz-Moreira et alibi., 2017). Water has dissolved O. The strain-feeding and single digestion system in tadpoles would increase the colonisation of facultatively anaerobic bacteria, such as Proteobacteria. For instance, Legionella pneumophilia, belonging to Proteobacteria, is common in many environments, including aquatic systems (Muraca et atomic number 13., 1990; Yu-Sen Maya Lin et alia., 1998). Rickettsiella, belonging to Proteobacteria, is also found in water and umpteen aquatic invertebrates (e.g., crustaceans) (Friedman et al., 1997; Küchler et al., 2009). In this study, both Legionella pneumophilia and Rickettsiella were significantly highest in abundance in MA than Megabyte, Mc, Oregon MD, where they were infrequent. In the MB stage (aquatic life style), the diet is the same as the MA stagecoach. However, the sexual climax of metamorphosis, including the increased complexity of the digestive arrangement, requires energy for ontogeny. At this stage, the tadpoles do not eat or eat less, and most of the energy for the metamorphosis operating room ontogenesis comes from the degeneration of tissues and obtained by oxidation of accumulated militia (e.g., fat and carbohydrate oxidisation) (Hourdry et al., 1996; Warne et al., 2017); these metabolic reactions will consume oxygen in the body. Thus, the hyperbolic complexity of the digestive system English hawthorn lead to a decrease in these facultatively anaerobic bacteria and an increase in anaerobic bacteria (e.g., Firmicutes).
Moreover, MC (terrestrial life style) is the windup of metabolism, including the complete biological process system. Therein step, most metabolic reactions by the host [e.g., degeneration of tails, corpulent, and carbohydrate (e.g., glycogen) oxidisation] and fermen by gut microbes go on to consume oxygen. Interestingly, Pass enrichment analysis at the MC stage (stage 45) showed that the majority of enriched GO categories were associated with saccharide metabolism (e.g., glycolysis, gluconeogenesis, hexose synthesis process, monosaccharide biosynthetic process, glucose catabolic summons, hexose catabolic process, monosaccharide biological process process, glucose metabolic process, single-organism saccharide catabolic process, carbohydrate catabolic treat, and carbohydrate synthesis process) (Zhao et al., 2016). Gut microbial function in the MC stage had the highest abundance in bacterium associated with sugar metabolic process among these four stages (Supplementary Physique S2). To boot, the new enzymes of the host related to food digestion and complexness of the digestive system (acidulent stomach and folded epithelial tissue of gut) further increase food fermentation and oxygen consumption in the gastrointestinal system. Thus, all these factors may bring to form the anaerobic home ground for the juvenile frog's gastrointestinal system in the MC stage and addition the colonization of anaerobiotic intestine microbes and the selection of particular intestine microbial groups together with the dietary shift. Considering the effect on the gut microbiome composition by luminal oxygen levels (He et al., 1999; Friedman et al., 2018), on that point is another affirmable example of the human relationship between the host and the catgut microbiome: the graduate carbohydrate metabolism level of intestine microbiomes in the MC stage whitethorn allow energy for host tissue paper rebuilding in addition to the vim provided by the host.
Additionally, in the MD (grown) stages, we constitute that the proportion of anaerobiotic bacterium decreased compared to the MC level. A piscivorous animal has a relatively simple and short systema digestorium (Hume, 2002). The adult frog has a large food ingestion (from large insects) compared to the juvenile salientia in the MC stages. Thus, the MD present will increase the commute of colonization by aerobic environmental bacteria, such as Verrucomicrobia, acquired from their new living conditions and even from their new diet. Verrucomicrobia is one of the dominant bacteria in the environment. The decrease in exploratory diversity of the gut microbiome from metamorphosis (herbivorous) to juvenile and mature frogs (insect-enriched) has besides been perceived between herbivorous and carnivorous vertebrates (Pasture et atomic number 13., 2008).
Conclusion
Here, we identified several putative relationships between the frog and its gut microbiome. (1) We revealed the catgut microbiome developmental association that was influenced by server dietary sources. (2) The remodeling of the gastrointestinal system during metamorphosis might besides have a profound effect along the gut microbial composition. (3) The possible functions of the gut microbiome could help the salientia's nutritional and energy necessarily during metabolism and evolution. Therefore, these findings provide the basic data for amphibious aircraft management and conservation.
Materials and Methods
Sample Collection and Gut Calm Preparation
A total of 80 singular tadpoles and 160 individual frogs (Auxiliary Table S1) were self-collected from the home ground close to the Noncivilised Research facility of the Chengdu Institute of Biology (E: 104°9′12, N: 31°6′35) located in Shifang County in Sichuan Province from June to August 2017. Based connected our premature experience on the feeding and maturation of M. fissipes in the captive surroundings and direct observation in the wild field [e.g., Zhao et al., (2016)], we could identify these tadpoles from M. fissipes and estimate the primary diet of M. fissipes. The tadpoles were self-contained lightly with fishing nets, and the young and adult individuals were captured after metamorphosis at night with unproductive gloves (usually 21:00 to 24:00 is the counteractive period). To see to it the unity of the table of contents in the alimentary tract, the frogs were immediately euthanized with MS-222. Tadpoles were baked at a assiduity of 0.3 to 0.6 g/L MS-222 for 2 to 3 min, and frogs were hardened for 5 to 8 Hokkianese or more. After washing the surface of the animals repeatedly with sterile distilled water to avoid residual solution, we removed the holonomic gastrointestinal tract from the abdominal cavity and scraped the inner wall of the intestines with sterile blades or squeezed the intestines directly away deal. All of the gut table of contents or biological process juices were placed into 2-mL centrifugal tubes and then stored at −80°C until DNA extraction. Because of the lack of enough gastrointestinal content in a single tadpole, subadult or even grownup individuals, we touch to the mixed treatments as the septrional Panthera pardus tadpoles and frogs (Kohl et al., 2013). In our study, four typical stages of growth and developmental process (Shimizu-Nishikawa et al., 2003; Wang et al., 2017) were selected: before forelimb and after hindlimb growth (S32–41), marked as radical MA; during the apex of metabolism and when the forelimb began to grow (S42–44), tagged every bit group MB; frogs that had late complete metamorphosis (S45), tagged every bit group Megacycle per second; and sexually ripen individuals, marked as group MD. In total, we obtained 18 qualified samples of intestinal contents or digestive juices, including four tubes in chemical group MA, four tubes in group MB, pentad tubes in group MC, and five tubes in group MD. Ten tadpole individuals were pooled for each tube (Tabular array 1).
Geomorphological and Histological Observations of Tadpole and Batrachian Intestines
We measured the mean ratio of the intestinal duration to body length among four groups (MA, 3; MB, 3; Megacycle per second, 3; and MD, 5 individuals). A stereo microscope (Nanjing Jiangnan Yongxin JSZ8 Stereo Microscope, Nanjing, China) with an MShot Digital Imaging System (microscope camera Mc50-N, Guangzhou, China) was wont to observe, take photos of tadpoles, and bar their trunk length and internal organ length. Then, the small intestines were dissected and 4% paraformaldehyde determinate. After dehydration in a graded serial of grain alcohol and foil past xylene, intestines were enclosed in paraffin and sectioned in serial publication transverse sections (4 μm thick) using an RM2016 [Leica RM2016 Manual Rotary Microtome (Wetzlar, Germany); Leica Microsystems]. Dewaxed serial sections were besmirched with Delafield's hematoxylin and counterstained with eosin to show general histologic characteristics.
DNA Extraction and Bacterial 16S rRNA Sequencing
Gastrointestinal samples were thawed on ice, and microbial genomic DNA was extracted using a QIAamp Fast Deoxyribonucleic acid Fecal matter Mini Kit (QIAGEN, Hilden, Germany) according to the manufacturer's protocol. The unity of the DNA was visually assessed using 1.0% agarose gel electrophoresis and quantified using a Qubit and NanoDrop. The highly versatile V4 region of the 16S rRNA gene was amplified from community genomic DNA exploitation the bacteria-specific world-wide primers 515F (GTGCCAGCMGCCGCGGTAA) and 806R (GGACTACHVGGGTWTCTAAT). Polymerase chain chemical reaction was performed in triplicate using a 25 μL reaction containing ∼40 ng of DNA template, 2.5 μL of 10 × TransStart Taq polisher, 1 μL of each forward and reverse ground, 2 μL of dNTPs (2.5 millimetre), 0.25 μL of TransStart Taq DNA Polymerase, and 16.25 μL of ddH2O. The polymerase chain reaction amplification conditions were A follows: initial denaturation at 94°C for 5 Min, followed by 35 cycles of denaturation at 94°C for 30 s, tempering at 53°C for 30 s, elongation at 72°C for 30 s, and a final university extension at 72°C for 10 min. Polymerase chain reaction products were pure with a Worldwide DNA Purification Kit (TIANGEN, Beijing, PRC), and barcoded V4 amplicons were sequenced using the Illumina HiSeq platform (HiSeq2500 PE250).
Information Analysis
Raw sequences were generated from the Illumina HiSeq sequencing program. We performed quality control (e.g., demultiplexing and denoising) and taxonomic group classification (founded on Silva 132 version) in QIIME1.9 (Caporaso et aliae., 2010). We obtained the usable taxonomic unit (OTU) abundance tables and diversity results for downstream analysis. We chose to rarefy our sequencing depth at ∼53,809 (reported to the lowest list of sequences of single sample in this subject) to equalize the sampling depth across complete samples.
Gut Microbic Community Psychoanalysis
We used LEfSe (Segata et al., 2011) to determine the gut microbial taxon with significantly differentiating abundance among groups (development stages). The relative abundance table of the bacteria families was inputted into Cytoscape 3.4.0 (Claude Elwood Shannon et atomic number 13., 2003). And then, we used the plugin CoNet (Faust and Raes, 2016) to generate co-occurrence plots victimisation these parameters (Spearman index, ρ = 0.7). CoNet can notice large not-unselected patterns of cobalt-happening (mutual excommunication and copresence) in abundance and relative incidence data (Faust and Raes, 2016). The alpha diversity (e.g., phylogenetic diversity) among groups was analyzed using one-way ANOVA in SPSS Statistics 20.0 (IBM SPSS, 2011).
Gut Microbial Genus Beta Diversity Analysis
The Bray-William Curtis aloofness for species abundance was used to bring forth NMDS in PAST3 (Hammer et aliae., 2001). Moreover, to evaluate the issue of growing stages on the gut microbiota composition, we performed one-way permutational multivariate Analysis of variance on Bray-Curtis dissimilarities in species copiousness in PAST3 (Hammer et alibi., 2001). To determine the dissimilarity trend concluded the development stages, we performed pairwise comparisons among groups on Unifrac unweighted distance (Lozupone et aliae., 2011). The contribution of each bowel microbic genus to the observed dissimilarity betwixt groups was performed by Similarity Percentages test (SIMPER) in PAST3 (Forge et atomic number 13., 2001).
Microorganism Phenotype Prediction
We victimized BugBase to predict the O-related phenotypes of the frog bowel microbiome (Montgomery Ward et alibi., 2017). BugBase is an algorithm that predicts being-level coverage of functional pathways also as biologically interpretable phenotypes so much as oxygen permissiveness and Gram staining within complex microbiomes using either marker gene sequencing data (e.g., 16S) or whole-genome scattergun data (Ward et al., 2017). For example, reference databases in BugBase enclosed Integrated Microbial Genomes (Markowitz et al., 2011), the Kyoto Cyclopaedia of Genes and Genomes (KEGG) (Kanehisa et al., 2011), and the Pathosystems Resource Integrating Core (Snyder et al., 2006), which were categorized to six major phenotypes (e.g., Gram staining, oxygen tolerance, ability to manikin biofilms, mobile element content, pathogenicity, and aerophilic stress tolerance) (Ward et al.., 2017). Here, the OTU table of 16S data was normalized by 16S written matter number then was stimulus into the BugBase to attain the trait prediction away algorithmic rule methods supported the extension databases (Ward et al., 2017). Pairwise Mann-Whitney-Wilcoxon tests were performed among groups.
Metagenomic Sequencing and Data Analysis
Metagenomic Sequencing
Metagenomic shotgun sequencing libraries were prepared and sequenced at Impress Biozeron Biological Technology Co. For for each one sample, 1 μg of genomic DNA was used with Illumina's TruSeq for program library preparation. Libraries were sequenced using the Illumina HiSeq 4000, PE 150.
Removal of Putative Host and Diet Contamination
HiSeq reads were filtered using custom Perl scripts and Trimmomatic (parameters: Trimmomatic-0.30.jounce PE -phred33 Stellar:0 Tracking:20 SLIDINGWINDOW:50:20 MINLEN:50) (Bolger et al., 2014) to remove (i) every last reads little than 50 bp in length, (ii) reads with degenerate bases (N′s), and (iii) all duplicates defined American Samoa sequences whose first 20 nucleotides were identical and shared an overall identity of greater than 97% throughout the length of the shortest read. After blasting with the NR databases using baseball field (Buchfink et al., 2014), we separate the putative host (frog) and diet (insect) contamination and gained just reads. This was the number one step to remove putative master of ceremonies contamination. Megahit (Li et AL., 2015) was accustomed assemble the clean reads (removing the contigs with coverage <60%), and prodigal was used for cistron anticipation (Hyatt et al., 2010). Then, we blasted these genes against the NR database in the National Center for Bioengineering Information using adamant and gained the putative taxon assignments of these genes per metagenome (Buchfink et alibi., 2014). Thus, we classified the taxon information for these genes into three categories, such atomic number 3 prokaryotes, and the different contamination sources (e.g., Host, diet). Then, we used salmon (Patro et atomic number 13., 2015) to map the clean reads to these genes per metagenome and kept exclusive the reads on paper belonging to prokaryotes. This was the second interfere removing the master of ceremonies or diet (insect) contamination per metagenome. We used megahit (Li et al., 2015) to assemble these clean reads into contigs and remove the contigs with insurance coverage of less than 60%. Prodigal (Hyatt et al., 2010) was used for cistron prevision from these peaky-calibre (flited) contigs, and we gained the gene files in this step. CD-HIT (Li and Godzik, 2006) was used to construct non-redundant gene sets with to a lesser degree 90% convergence and to a lesser degree 95% shared sequence indistinguishability from these gene files. We exploited diamond (Buchfink et al., 2014) to conduct species annotation for the non-redundant gene profile, remove the genes putatively belonging to the host and dieting, and gain the final clean not-redundant gene profile. This was the closing step to remove contamination. The referred publicized batrachian genome is non considerably made-up, which pass to the poor blaring for some metagenome reads. Thus, we used this ordinal step to remove the acknowledged contaminations based on non-redundant gene sets.
Pure Read Processing and KEGG Analysis
Based on these gene profiles, we used chromatic (Patro et al., 2015) to map the clean reads (from step 2 aft removing contamination) per metagenome to the clean non-redundant gene profile and determine the TPM abundance (transcripts per million reads) of these not-surplus gene profiles in each metagenome. Finally, the clean not-spare gene sequences were searched against the KEGG database using diamond (Buchfink et al., 2014). The KEGG orthology, enzyme commission, and KEGG pathways associated with each sequence were determined. We calculated the relative abundance of KEGG pathways using TPM (transcripts per million) (Wagner et al., 2012).
Transcriptome Analysis
We reanalyzed the transcriptome information in our previously published study (Zhao et Heart of Dixie., 2016). These transcriptomes represented the three key biological process stages of M. fissipes: premetamorphosis (stage 30), hemimetabolous climax (stage 42, MB), and completion of metamorphosis (stage 45, MC). Three individuals from for each one of the three stages were used as individual natural replicates. Here, we focused on the genes cryptography for putative enzymes involved in the chitin metabolic pathway collect to the fare changes in these frogs. To compare the unigene expression levels, each unigene was further normalized by fragments per kilobase of coding DNA model per million mapped reads for the three organic process stages (Trapnell et al., 2010). The transcriptome analytic thinking focused on the changes in factor expression during metabolism and did not include the full-grown arrange samples.
Author's Note
This study has also used the old published transcriptome data (Zhao et al., 2016) with accession issue (GECV00000000) at NCBI.
Data Accessibility Statement
The 16S sequencing data and mapfile have been deposited in figshare (10.6084/m9.figshare.11627460). The natural metagenome data is organism uploaded to NCBI (National Center for Biotechnology Data) with the accession identification number PRJNA601807. The requirement of the crude data and materials in this study can also send to LZ (zhulf@ioz.ac.cn).
Ethics Statement
The foxlike wont protocol in that study (permit: 2015-Argon-JJP-01) was reviewed and approved by the Animal Ethical and Welfare Citizens committee of Chengdu Institute of Biology, Chinese Academy of Sciences, Chengdu, People's Republic of China. The Chairman of this committee is Dr. Xinquan Zhao.
Author Contributions
LZ and JJ conceived the project. MZ, LX, XW, and LC accumulated the samples. MZ performed the experiments. LZ, MZ, HC, LL, and GL analyzed the data. MZ, QC, LZ, JJ, and LG wrote the manuscript. All authors gave final approval for publication.
Funding
This search was financially pendant by the Status Key Program of Search and Evolution, Ministry of Skill and Technology (2016YFC0503200), Important Research Project of the Chinese Academy of Sciences (KJZG-EW-L13), National Natural Scientific Funds (31491964) to JP, Sichuan Science and Technology Project (2017JY0339), West Lightsome Institution of the Chinese Academy of Sciences (2015XBZG_XBQNXZ_B_011) to LL, and the Priority Academic Syllabu Evolution of Jiangsu High Education Institutions (PAPD).
Struggle of Interest
HC was employed by Hangzhou Legenomics Bio-Pham Technology Atomic number 27., Ltd.
The remaining authors declare that the research was conducted in the petit mal epilepsy of any commercial or financial relationships that could embody construed A a potential conflict of interest.
Acknowledgments
We give thanks Wiley Editing Services for the language editing. We thank Guocheng Shu, Shouhong Wang, Wei Zhu, and Xiaoyi Wang for their help and suggestions during the sample collection and homework of the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2020.00162/full#supplementary-material
References
Barber, C. B., Dobkin, D. P., and Huhdanpaa, H. (1996). The quickhull algorithm for convex hulls. ACM Trans. Math. Softw. 22, 469–483. Interior Department: 10.1145/235815.235821
CrossRef Full-of-the-moon Text | Google Scholar
Caporaso, J. G., Kuczynski, J., Stombaugh, J., Bittinger, K., Bushman, F. D., Costello, E. K., et al. (2010). QIIME allows analysis of high-throughput biotic community sequencing data. Nat. Methods 7, 335–336.
Google Scholar
Castaneda, L. E., Sabat, P., Gonzalez, S. P., and Nespolo, R. F. (2006). Biological process plasticity in tadpoles of the Chilean giant frog (Caudiverbera caudiverbera): factorial effects of diet and temperature. Physiol. Biochem. Zool. 79, 919–926. doi: 10.1086/506006
PubMed Abstract | CrossRef Full Text | Google Student
Chai, L., Dong, Z., Chen, A., and Wang, H. (2018). Changes in enteric microbiota of Bufo gargarizans and its connexion with body weight during metamorphosis. Arch. Microbiol. 200, 1087–1099. Interior Department: 10.1007/s00203-018-1523-1
PubMed Abstract | CrossRef Full Text | Google Scholar
Chen, B., Teh, B.-S., Sunbathe, C., Hu, S., Lu, X., Boland, W., et al. (2016). Biodiversity and activenes of the bowel microbiota crossways the life history of the insect herbivore Spodoptera littoralis. Sci. Repp. 6:29505. doi: 10.1038/srep29505
PubMed Abstract | CrossRef Full Text | Google Scholar
Daniel, H., Gholami, A. M., Berry, D., Desmarchelier, C., Hahne, H., Loh, G., et al. (2014). Postgraduate-fat diet alters gut microbiota physiology in mice. ISME J. 8, 295–308. doi: 10.1038/ismej.2013.155
PubMed Abstract | CrossRef Full Text | Google Scholar
Dodd, M., and Dodd, J. (1976). The biology of metamorphosis. Physiol. Amphib. 3, 467–599.
Google Scholar
Doucet, D., and Retnakaran, A. (2012). Dirt ball chitin: metastasis, genomics and gadfly management. Adv. In Insect Phys. 43, 437–511.
Google Scholar
Fisher, R. (1955). The absorption of water and of some small solute molecules from the isolated small intestine of the rat. J. Physiol. 130, 655–664. doi: 10.1113/jphysiol.1955.sp005433
CrossRef Total Text edition | Google Scholarly person
Friedman, C. S., Thomson, M., Chun, C., Haaker, P. L., and Hedrick, R. P. (1997). Withering syndrome of the black-market abalone, Genus Haliotis cracherodii (Leach): water temperature, intellectual nourishment availability, and parasites As possible causes. J. Shellfish Reticuloendothelial system. 16, 403–411.
Google Scholar
Milton Friedman, E. S., Bittinger, K., Esipova, T. V., Hou, L., Chau, L., Jiang, J., et al. (2018). Microbes vs. chemistry in the parentage of the anaerobiotic gut lumen. Proc. Natl. Acad. Sci. AmericaA. 115, 4170–4175. doi: 10.1073/pnas.1718635115
PubMed Ideational | CrossRef Full Text | Google Assimilator
Gould, A. L., Zhang, V., Lamberti, L., Jones, E. W., Obadia, B., and Korasidis, N. (2018). Microbiome interactions shape Host fitness. Proc. Natl. Acad. Sci. United States of AmericaA. 115, E11951–E11960. doi: 10.1073/pnas.1809349115
PubMed Cabbage | CrossRef Chockful Textual matter | Google Scholar
He, G., Shankar, R. A., Chzhan, M., Samouilov, A., Kuppusamy, P., and Zweier, J. L. (1999). Noninvasive measurement of physical structure structure and intraluminal oxygenation in the gastrointestinal tract of bread and butter mice with spatial and array EPR imaging. Proc. Natl. Acad. Sci. U.S.A. 96, 4586–4591. doi: 10.1073/pnas.96.8.4586
PubMed Abstract | CrossRef Full Textbook | Google Scholar
Hendricks, F. S. (1973). Intestinal contents of Rana pipiens Schreber (Ranidae) larvae. Southwest. Nat. 18, 99–101.
Google Scholar
Hoefel, D., Monis, P. T., Grooby, W. L., Andrews, S., and Saint, C. P. (2005). Polish-indie techniques for rapid spying of bacteria connected with loss of chloramine residual in a drinking water. Appl. Encircle. Microbiol. 71, 6479–6488. Interior Department: 10.1128/aem.71.11.6479-6488.2005
PubMed Abstract | CrossRef Full Text | Google Scholar
Hourdry, J., L'hermite, A., and Ferrand, R. (1996). Changes in the digestive nerve tract and feeding behavior of anuran amphibians during metamorphosis. Physiol. Zool. 69, 219–251. Department of the Interior: 10.1086/physzool.69.2.30164181
CrossRef Chock-full Text | Google Scholar
Hume, I. D. (2002). Digestive strategies of mammals. Acta Zool. Sin. 48, 1–19.
Google Scholar
Hyatt, D., Chen, G.-L., Locascio, P. F., State, M. L., Larimer, F. W., and Hauser, L. J. (2010). Unstinted: prokaryotic factor realization and translation initiation web site recognition. BMC Bioinformatics 11:119. doi: 10.1186/1471-2105-11-119
PubMed Synopsis | CrossRef Engorged Text | Google Scholar
IBM SPSS (2011). IBM SPSS Statistics for Windows, Version 20.0. Unused York, NY: IBM Corp.
Google Scholar
Ishizuyaoka, A. (2011). Amphibious aircraft organ remodeling during metamorphosis: insight into thyroid hormone−iatrogenic apoptosis. Dev. Growth Dissent. 53, 202–212. doi: 10.1111/j.1440-169X.2010.01222.x
PubMed Notional | CrossRef Full Text | Google Scholar
Kaakoush, N. O. (2015). Insights into the role of Erysipelotrichaceae in the human host. Front. Cell. Taint. Microbiol. 5:84. Interior: 10.3389/fcimb.2015.00084
CrossRef Full Text | Google Scholar
Kanehisa, M., Goto, S., Sato, Y., Furumichi, M., and Tanabe, M. (2011). KEGG for integrating and interpretation of large-scale molecular data sets. Nucleic Acids Res. 40, D109–D114. doi: 10.1093/nar/gkr988
PubMed Abstract | CrossRef Full Text | Google Scholar
Kohl, K. D., Cary, T. L., Karasov, W. H., and Dearing, M. D. (2013). Restructuring of the amphibian bowel microbiota finished transfiguration. Environ. Microbiol. Rep. 5, 899–903. doi: 10.1111/1758-2229.12092
PubMed Abstract | CrossRef Full Textual matter | Google Learner
Koliada, A., Syzenko, G., Moseiko, V., Budovska, L., Puchkov, K., Perederiy, V., et al. (2017). Association between body mass index finger and Firmicutes/Bacteroidetes ratio in an adult Ukrainian population. BMC Microbiol. 17:120. doi: 10.1186/s12866-017-1027-1
PubMed Abstract | CrossRef Full Text | Google Scholar
Kramer, K. J., and Muthukrishnan, S. (1997). Insect chitinases: unit biological science and potential purpose as biopesticides. Dirt ball Biochem. Mol. Biol. 27, 887–900. doi: 10.1016/s0965-1748(97)00078-7
PubMed Nonobjective | CrossRef Full Text | Google Scholar
Küchler, S. M., Kehl, S., and Dettner, K. (2009). Characterisation and localization of Rickettsia sp. in water beetles of genus Deronectes (Coleoptera: Dytiscidae). FEMS Microbiol. Ecol. 68, 201–211. doi: 10.1111/j.1574-6941.2009.00665.x
PubMed Nonfigurative | CrossRef Full Text | Google Scholar
Laudet, V. (2011). The origins and evolution of vertebrate metamorphosis. Curr. Biol. 21, R726–R737.
Google Scholar
Lecomte, V., Kaakoush, N. O., Maloney, C. A., Raipuria, M., Huinao, K. D., Mitchell, H. M., et al. (2015). Changes in gut microbiota in rats federal official a pinched fat dieting correlate with obesity-associated organic process parameters. PLoS One 10:e0126931. Interior Department: 10.1371/journal.pone.0126931
PubMed Nonobjective | CrossRef Full Text | Google Scholar
Le, R. E., Bäckhed, F., Turnbaugh, P., Lozupone, C. A., Knight, R. D., and Gordon, J. I. (2005). Obesity alters catgut microbial ecology. Proc. Natl. Acad. Sci. AmericaA. 102, 11070–11075.
PubMed Abstractionist | Google Scholar
Ley, R. E., Lozupone, C. A., Hamady, M., Knight, R., and Gordon, J. I. (2008). Worlds within worlds: evolution of the vertebrate gut microbiota. Nat. Rev. Microbiol. 6, 776–788. doi: 10.1038/nrmicro1978
PubMed Synopsis | CrossRef Full Text | Google Scholarly person
Pasture, R. E., Turnbaugh, P. J., Klein, S., and Gordon, I. (2006). Microbial ecology: human gut microbes associated with obesity. Nature 444, 1022–1023.
PubMed Abstract | Google Scholar
Li, D., Liu, C.-M., Luo, R., Sadakane, K., and Lam, T.-W. (2015). MEGAHIT: an ultra-fast one-node result for large and hard metagenomics assembly via succinct Delaware Bruijn graph. Bioinformatics 31, 1674–1676. doi: 10.1093/bioinformatics/btv033
PubMed Abstract | CrossRef Full Text | Google Learner
Li, W., and Godzik, A. (2006). Cd-hit: a fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 22, 1658–1659. DoI: 10.1093/bioinformatics/btl158
PubMed Nonfigurative | CrossRef Full Text | Google Scholar
Linzey, D. W. (1967). Solid food of the leopard frog, Rana p. pipiens, in central Unprecedented House of York. Herpetologica 23, 11–17.
Google Scholar
Lozupone, C., Lladser, M. E., Knights, D., Stombaugh, J., and Knight, R. (2011). UniFrac: an effective distance metric for microbial community comparison. ISME J. 5, 169–172. Interior Department: 10.1038/ismej.2010.133
PubMed Abstract | CrossRef Full Textbook | Google Scholar
Markowitz, V. M., Chen, I.-M. A., Palaniappan, K., Chu, K., Szeto, E., Grechkin, Y., et aluminium. (2011). IMG: the integrated micro-organism genomes database and comparative analysis system. Nucleic Acids RES. 40, D115–D122.
Google Scholar
Martínez, I., Perdicaro, D. J., Brown, A. W., Hammons, S., Carden, T. J., and Carr, T. P. (2012). Diet-elicited alterations of host cholesterol metabolism are probable to affect gut microbiota composition in hamsters. Appl. Encircle. Microbiol. 79, 516–524. doi: 10.1128/AEM.03046-12
PubMed Ideal | CrossRef Full Text | Google Scholar
McFall-Ngai, M., Hadfield, M. G., Bosch, T. C., Carey, H. V., Domazet-Lošo, T., Douglas, A. E., et al. (2013). Animals in a microorganism world, a spick-and-span imperative for the life sciences. Proc. Natl. Acad. Sci. U.S.A. 110, 3229–3236. doi: 10.1073/pnas.1218525110
PubMed Purloin | CrossRef Full Text | Google Learner
Muraca, P. W., Victor, L. Y., and Goetz, A. (1990). Disinfection of water system statistical distribution systems for Legionella pneumophilia: a review of application procedures and methodologies. Infect. Ascendancy Hosp. Epidemiol. 11, 79–88. doi: 10.1086/646126
PubMed Abstract | CrossRef Egg-filled Text | Google Scholar
Nicholson, J. K., Holmes, E., Kinross, J., Burcelin, R., Charles Dana Gibson, G., Jia, W., et Alabama. (2012). Host-gut microbiota metabolous interactions. Science 336, 1262–1267. doi: 10.1126/science.1223813
PubMed Abstract | CrossRef Full Text | Google Scholar
Patro, R., Duggal, G., and Kingsford, C. (2015). Salmon: right, versatile and ultrafast quantification from RNA-seq data victimization whippersnapper-alignment. bioRxiv [Preprint]. doi: 10.1101/021592
CrossRef Full Textbook | Google Scholar
Schreiber, A. M., Cai, L., and Brown, D. D. (2005). Remodeling of the intestine during metamorphosis of Xenopus laevis. Proc. Natl. Acad. Sci. U.S.A. 102, 3720–3725. doi: 10.1073/pnas.0409868102
PubMed Conceptional | CrossRef Full Text | Google Scholar
Segata, N., Izard, J., Waldron, L., Gevers, D., Miropolsky, L., Garrett, W. S., et al. (2011). Metagenomic biomarker discovery and explanation. Genome Biol. 12:R60. Interior: 10.1186/gb-2011-12-6-r60
PubMed Abstract | CrossRef Full Textbook | Google Bookman
Claude Shannon, P., Markiel, A., Ozier, O., Baliga, N. S., Wang, J. T., Ramage, D., et al. (2003). Cytoscape: a software environment for co-ordinated models of biomolecular fundamental interaction networks. Genome Res. 13, 2498–2504. doi: 10.1101/gr.1239303
PubMed Abstract | CrossRef Chuck-full Text | Google Scholar
Shi, Y.-B. (2000). Amphibious vehicle Transfiguration. NY, New York State: Wiley-Liss.
Google Scholarly person
Shimizu-Nishikawa, K., Takahashi, J., and Nishikawa, A. (2003). Intercalary and unneeded regeneration in the limbs of the frog, African clawed frog. Dev. Dyn. 227, 563–572. doi: 10.1002/dvdy.10345
PubMed Abstract | CrossRef Full Text | Google Scholar
Adam Smith, D. C., and Douglas, A. E. (1987). The Biology of Symbiosis. London: Black Prince Arnold (Publishers) Ltd.
Google Scholar
Snyder, E., Kampanya, N., Lutecium, J., Nordberg, E. K., Karur, H., Shukla, M., et aliae. (2006). PATRIC: the VBI pathosystems resourcefulness desegregation center. Nucleic Acids Res. 35, D401–D406.
PubMed Abstract | Google Scholar
Suzuki, M., Fujimoto, W., Goto, M., Morimatsu, M., Syuto, B., and Iwanaga, T. (2002). Cellular expression of bowel chitinase mRNA in the gastrointestinal tract of mice and chickens. J. Histochem. Cytochem. 50, 1081–1089. doi: 10.1177/002215540205000810
PubMed Abstract | CrossRef Wide Textual matter | Google Scholar
Tetlock, A., Yost, C. K., Stavrinides, J., and Manzon, R. G. (2012). Changes in the gut microbiome of the sea lamprey during metabolism. Appl. Environ. Microbiol. 78, 7638–7644. doi: 10.1128/AEM.01640-12
PubMed Abstract | CrossRef Full Text | Google Student
Count Rumford, S. N. (2003). Trehalose – the insect 'Rip' dough. Adv. In Insect Physiol. 31, 205–285. doi: 10.1016/s0065-2806(03)31004-5
CrossRef Filled Text | Google Scholar
Trapnell, C., Theodore Samuel Williams, B. A., Pertea, G., Mortazavi, A., Kwan, G., Avant-garde Baren, M. J., et Heart of Dixie. (2010). Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 28, 511–515. doi: 10.1038/nbt.1621
PubMed Abstract | CrossRef Cram full Text | Google Learner
Turnbaugh, P. J., Bäckhed, F., Robert Fulton, L., and Gordon, J. I. (2008). Diet-iatrogenic corpulency is linked to marked but reversible alterations in the mouse distal gut microbiome. Cell Host Microbe 3, 213–223. doi: 10.1016/j.chom.2008.02.015
PubMed Filch | CrossRef Engorged Text | Google Scholar
Turnbaugh, P. J., Ley, R. E., Mahowald, M. A., Magrini, V., Mardis, E. R., and Gordon, I. (2006). An obesity-associated catgut microbiome with increased capacity for DOE harvest. Nature 444, 1027–1031.
PubMed Abstract | Google Student
Turnbaugh, P. J., Ridaura, V. K., Faith, J. J., Rey, F. E., Knight, R., and Gordon, I. (2009). The effect of diet on the human intestine microbiome: a metagenomic analysis in humanized gnotobiotic mice. Sci. Transl. Med. 1, ra14–ra16. Department of the Interior: 10.1126/scitranslmed.3000322
PubMed Abstract | CrossRef Full Textbook | Google Learner
Vaz-Moreira, I., Nunes, O. C., and Manaia, C. M. (2017). Omnipresent and unforgettable Proteobacteria and other Gram-negative bacteria in drinking weewe. Sci. Total Environ. 586, 1141–1149. doi: 10.1016/j.scitotenv.2017.02.104
PubMed Pilfer | CrossRef Full Text | Google Scholar
Verdam, F. J., Fuentes, S., Diamond State Jonge, C., Zoetendal, E. G., Erbil, R., Greve, J. W., et aliae. (2013). Human enteral microbiota composition is associated with local and systemic inflammation in corpulency. Obesity 21, E607–E615. doi: 10.1002/oby.20466
PubMed Abstract | CrossRef Full Text | Google Scholar
Wagner, G. P., Kin, K., and Lynch, V. J. (2012). Measurement of mRNA abundance using RNA-seq data: RPKM measure is inconsistent among samples. Hypothesis Biosci. 131, 281–285. doi: 10.1007/s12064-012-0162-3
PubMed Abstract | CrossRef Full Text | Google Scholarly person
Wang, S., Zhao, L., Liu, L., Yang, D., Khatiwada, J. R., Wang, B., et aluminum. (2017). A stark embryonic developmental shelve of Microhyla fissipes (Amphibia, Salientia, Brevicipitidae). Asian Herpetol. Res. 8, 108–117.
Google Scholar
Ward, T., Larson, J., Meulemans, J., Hillmann, B., Lynch, J., and Sidiropoulos, D. (2017). BugBase predicts organism level microbiome phenotypes. bioRxiv [Preprint]. doi: 10.1101/133462
CrossRef Full Text | Google Scholarly person
Warne, R. W., Kirschman, L., and Zeglin, L. (2019). Manipulation of gut microbiota during critical developmental Windows affects host physiological performance and disease susceptibility across ontogeny. J. Anim. Ecol. 88, 845–856. doi: 10.1111/1365-2656.12973
PubMed Abstract | CrossRef Full Text | Google Scholar
Warne, R. W., Kirschman, L. J., and Zeglin, L. H. (2017). Manipulation of gut microbiota reveals unsteady community structure shaped away master of ceremonies biological process windows in Class Larvae. Integr. Comp. Biol. 57, 786–794. doi: 10.1093/icb/icx100
PubMed Abstract | CrossRef Full Text | Google Scholar
Werner, E. E. (1986). Amphibian metamorphosis: rate of growth, depredation risk, and the optimal size at transformation. Am. Nat. 128, 319–341. doi: 10.1086/284565
CrossRef Full Text | Google Scholar
Wilbur, H. M., and Collins, J. P. (1973). Ecological aspects of class metamorphosis: nonnormal distributions of competitive ability reflect selection for facultative transfiguration. Science 182, 1305–1314. doi: 10.1126/skill.182.4119.1305
PubMed Abstract | CrossRef Booming Text | Google Scholar
Wlodarska, M., Kostic, A. D., and Xavier, R. J. (2015). An centralising view of microbiome-host interactions in inflammatory intestine diseases. Cell Host Bug 17, 577–591. doi: 10.1016/j.chom.2015.04.008
PubMed Abstract | CrossRef Full School tex | Google Scholar
Yatsunenko, T., Rey, F. E., Manary, M. J., Trehan, I., Dominguez-Bello, M. G., and Contreras, M. (2012). Homo catgut microbiome viewed crossways eld and geography. Nature 486, 222–227. doi: 10.1038/nature11053
PubMed Impalpable | CrossRef Full Text | Google Student
Yu-Sen Lin, E., Fat, J. E., Yu, V. L., and Vidic, R. (1998). Disinfection of water distribution systems for Legionella. Semin. Respir. Infect. 13, 147–159.
Google Scholar
Zhao, L., Liu, L., Wang, S., Wang, H., and Jiang, J. (2016). Transcriptome profiles of metamorphosis in the ornamented pygmy frog Microhyla fissipes clear up the functions of thyroid hormone receptors in metamorphosis. Sci. Repp. 6:27310. doi: 10.1038/srep27310
PubMed Abstract | CrossRef Total Text | Google Scholar
Zhu, L., Wu, Q., Dai, J., Zhang, S., and Wei dynasty, F. (2011). Tell of cellulose metabolism by the giant panda gut microbiome. Proc. Natl. Acad. Sci. U.S.A. 108, 17714–17719. doi: 10.1073/pnas.1017956108
PubMed Abstract | CrossRef Full Text | Google Scholar
how long is the small intestine in a frog
Source: https://www.frontiersin.org/articles/10.3389/fmicb.2020.00162/full

{kind=link}
Posting Komentar untuk "how long is the small intestine in a frog"